
Grid Cells in rat entorhinal cortex were discovered in the Moser lab in Trondheim, Norway. These cells were first described in a paper in Nature 20051; For the past 8 years these neurons have been objects of intense study. As the New York Times reports, a paper published yesterday in Nature Neuroscience2 indicates rats aren't the only animals with grid cells; people have them, too. What are grid cells and what is the significance of recording them in humans?
Grid Cells, Place Cells and Navigation. Understanding grid cells requires a fair amount of background and context.
Navigation. From the NY Times article:
Scientists’ discovery that rodents, bats and nonhuman primates have a system in the brain for what amounts to dead reckoning navigation...
The 'dead reckoning' system referred to is the grid cell system. Dead reckoning refers to the ability to navigate without external cues. The term comes from ship navigation, where a ship will "take a sighting" via cues such as the stars or landmarks to determine where the ship is on a map, and then, as the ship moves, 'dead reckon' to update location on the map paying attention to speed and direction. Keeping track of location on a map is a combination of taking sightings and dead reckoning. For us, 'dead reckoning' refers to an ability to keep track of our position in space as we move with eyes closed or not attending to landmarks. The general notion is that a human or animal updates it's location on an internal map by a mix of 'dead recking' and 'sightings'. The hippocampus and entorhinal cortex contribute to both of these functions.
Grid Cells and Place Cells are types of neurons recorded from neighboring forebrain structures — grid cells from entorhinal cortex, place cells from the hippocampus. Each of these cell types was discovered in recordings from freely behaving rats. The rats must be unrestricted because viewing the defining firing patterns requires that the rat roam across a large apparatus.
Recording Procedure. Place cells and grid cells cannot be understood without an appreciation of the recording procedure. A few weeks prior to an experiment a rat has an electrode bundle surgically implanted into the target brain region and a connector for the electrodes is cemented on the head. After recovery, a connecting cable is attached daily and the electrodes are scanned in search of 'good cells'. If an electrode tip is sufficiently close to a neuron it can record the neuron's spikes (action potentials). When one or more electrodes are successfully positioned, experiments are run. During an experiment spike trains are recorded in parallel with x,y coordinates of the rat's location as viewed from above. (A small light fixed to the rat's head aids in tracking.) The figure below illustrates the technique for recording and depicting place-cell firing patterns; aside from electrode placement, the technique for grid cells is identical. The figure is from work I did with Bob Muller and Jim Ranck in the 1980s4; the basic procedures have not changed.
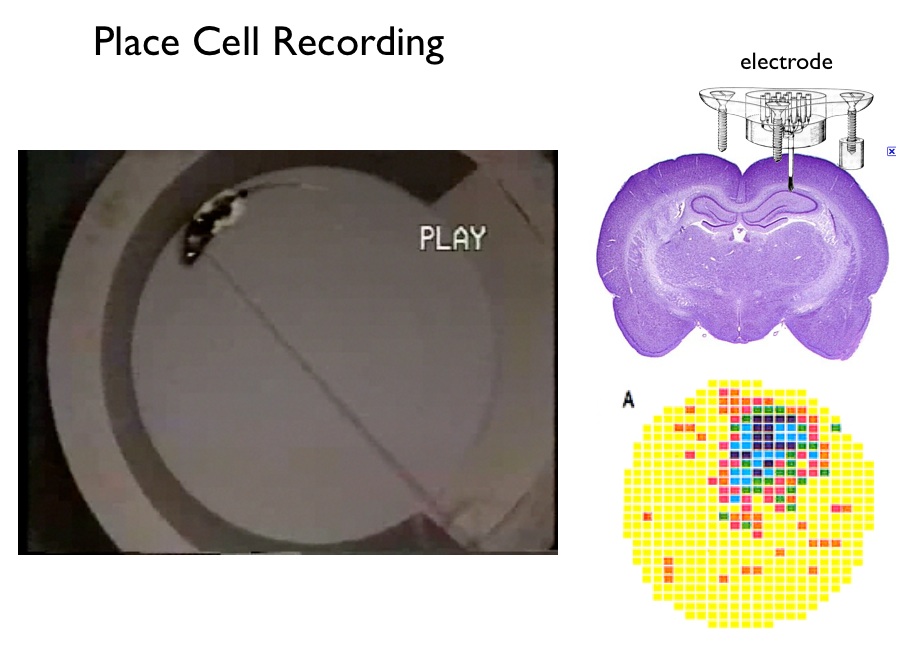
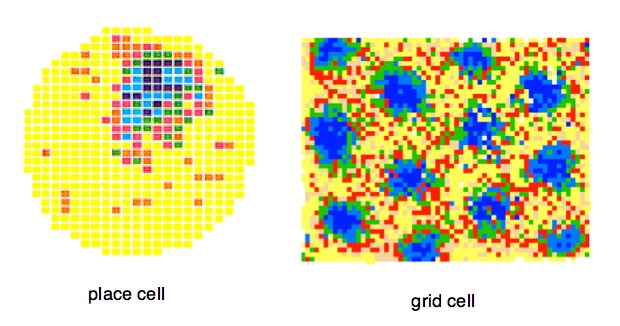
Place Cells. As illustrated above, a typical hippocampal place cell fires in a single region in the apparatus — this is the cell's 'firing field'. Place cells were discovered by John O'Keefe of University College, London in the early 1970s. John O'Keefe and Lynn Nadel suggested that thousands of place cells, covering the surface of any space, acted as a mapping system, and suggested that the hippocampus as a whole created a "cognitive map". In support is the general finding after hippocampal damage rats are terrible at spatial problem solving and appear lost in space. The general sense is that the firing of each place cell signals recognition of a specific place in a familiar environment.
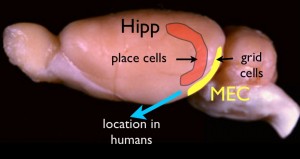
Grid Cells were discovered roughly 30 years later in the laboratory of Edvard and May-Britt Moser in Trondheim, Norway1. Grid cells are found in the entorhinal cortex, a region on the posterior wall of the rat's cerebral hemisphere. In the figure to the right, the entorhinal cortex is depicted in yellow. (this is a lateral view; viewed from the back EC is larger and pancake-shaped). The grid-cell and place-cell maps in the figure below were made with identical recording and analytic methods. In some ways the patterns of the two cell types are similar: in each the spatial signal is strong — that is, each exhibits strong "location-specific firing". But the grid cell pattern is different. Rather than a single 'firing field' in the apparatus, there are many fields (or 'bumps'). Additionally, the firing fields form a regular pattern of triangles (or hexagons). The signal from a single grid cell is complex. It cannot be a place recognition signal, because there are many places. The firing of a grid cell (on a bump) signals the distance and angle to other firing locations. Grid cell firing appears to be a signal that can be used for measuring displacement distances and direction. In other words, a 'metric'.
Modular Organization of Grid Cells Within a local regions of entorhinal cortex all grid cells have the same bump spacing (scale) and orientation (angle of the rows and columns), but differ in bump placement.The local regions are called 'modules'. One can think of all of the cells in module sharing a single bump pattern that can be moved horizontally or vertically, with a different x- y- offset for each cell. This means that as the rat moves, different grid cells in the module are excited. The distance and direction of movement determines the specific sequence of grid cells that will fire. Conversely, the sequence of activated grid cells signals speed, distance and direction of movement. This is thought to be the grid cell code. The figure below illustrates. Each set of colored circles represents the firing pattern of one grid cell. Notice the hexagonal arrangement for the circles of any one color. All illustrated cells are in single module. Now imagine the rat is at sitting at a location of one of the bumps for grid cell A. Now it moves south-west a certain distance. Grid cells B and C will fire. This identical firing pattern will occur whenever the rat is at position A and moves SW, In other words, Firing A, B, C codes movement from A southwest, independent of which A bump we start with. The output of a grid cell modules codes movement distance and direction.
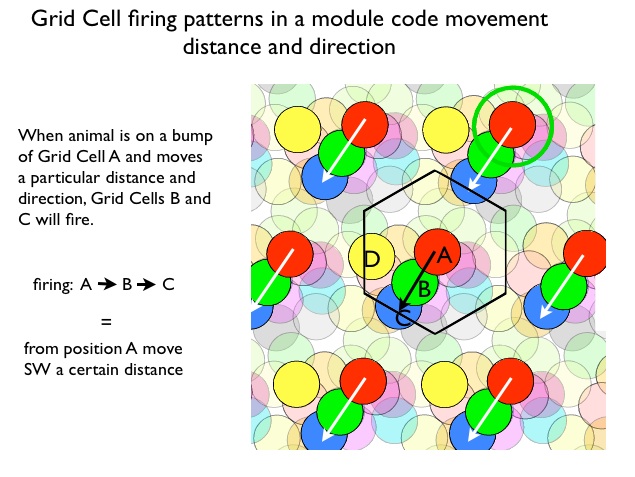
This shows why the the grid cell system is an excellent candidate for computing dead reckoning. As the rat moves, the grid cell system is consistently updating distance and direction.
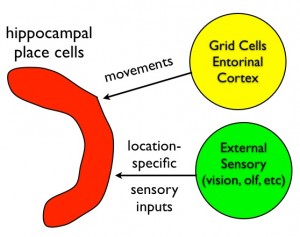
The emerging idea is as follows. Hippocampal place cells behave as a navigator on the ocean, updating the estimate of location using two types of inputs: dead reckoning and sightings. The grid cell system described above is thought to provide the path integration (dead reckoning) input. Other pathways into the hippocampus (probably lateral entorhinal cortex) convey sensory information for "sighting". The collection of hippocampal place cells combine both types of inputs to create a map. Map processing identifies familiar locations and computes efficient paths to other locations.
Learning. The story is not finished. Neither rats nor humans are born with knowledge of the layout of any environment — specific features of each environment must be learned. The general notion is that the grid cell system plays little direct role in learning; the "learning" (plasticity) lies in the synapses onto place cells that can change strength through experience. In brief, place cells are thought to be the location of binding between sensory cues and specific locations, while grid cells tie the map together signaling distances and directions connecting locations. The conjunction of place cells and grid cells forms a map in which an animal can recognize when it is in a familiar location in a familiar environment, and compute efficient paths that will take it to other familiar locations.
About the Author

I work at Downstate Medical Center in Brooklyn, NY, where my time is divided between research, teaching and avoiding committees. In grad school I studied olfaction in snakes and salamanders, but I’ve been studying the rat hippocampus and its relations to navigation and memory since the 1980s.
References
--- Resources:
This past spring the New York Times ran a profile of the Mosers with interesting links on grid cells: How Grid Cells in Brain Help Map out Space. A sense of where you are (profile with video). Crystals in the Brain. Article for general audiences written by the Mosers.
 Place cell video from Matt Wilson's lab. Current techniques permit recordings of 10s of neurons simultaneously. Perhaps up to 100. In the video spikes are colored dots, each place cell is a different color. (click on image to go to youtube)
Place cell video from Matt Wilson's lab. Current techniques permit recordings of 10s of neurons simultaneously. Perhaps up to 100. In the video spikes are colored dots, each place cell is a different color. (click on image to go to youtube)
---- References and Notes
1Hafting, T., Fyhn, M., Molden, S., Moser, M.-B., & Moser, E. I. (2005). Microstructure of a spatial map in the entorhinal cortex. Nature, 436(7052), 801–806. doi:10.1038/nature03721
2Jacobs, J., Weidemann, C. T., Miller, J. F., Solway, A., Burke, J. F., Wei, X.-X., et al. (2013). Direct recordings of grid-like neuronal activity in human spatial navigation. Nature Neuroscience. doi:10.1038/nn.3466. Published online Aug 4, 2013
3O'Keefe, J., & Nadel, L. (2008). The Hippocampus as a Cognitive Map - Oxford Press 1978, 296. full text online.
4Muller, R. U., Kubie, J. L., & Ranck, J. B. (1987). Spatial firing patterns of hippocampal complex-spike cells in a fixed environment. The Journal of neuroscience, 7(7), 1935–1950.
5It should be noted that this is not the first evidence of a grid cell system in humans. Deoller et al, (2010) using fMRI techniques reported strongly suggestive evidence. But Doeller et al did not record directly from grid cells. (Doeller, C. F., Barry, C., & Burgess, N. (2010). Evidence for grid cells in a human memory network. Nature, 463(7281), 657–661. doi:10.1038/nature08704)
6 Tiles have special properties. A "tile" is shape that contains one and only one bump in a module. Several shapes work: rectangles, parallelograms and a hexagon. A tile can be spread evenly ("tiled") across the floor of the apparatus, each defining a repeat of the module.
Additionally, a tile has no special 'center'; it can be translated horizontally or vertically and still contain one bump for each grid cell.
Also In Anatomy






Trending
Popular articles on BrainFacts.org








